
نوع مقاله : مقاله پژوهشی
Introduction
Adipose tissue is an active endocrine organ that secretes bioactive molecules called adipokines that participate in the regulation of metabolic homeostasis. Some studies have reported that in addition to nutrition, some genes have an effect on the growth and proliferation of subcutaneous adipose tissue. One of these genes is transcription factor 4 (TCF4). Mutations in TCF4 can cause digestive problems that may result in the growth and proliferation of subcutaneous fat cells. Some genes act as an antagonist of TCF4 and prevent the reproduction and transcription of this gene. One of these genes is CHOP10 (C/EBP homologous protein). It is a basic-leucine zipper protein that plays a role in differentiation and apoptosis. The CHOP10 is involved in a variety of stress response pathways, including endoplasmic reticulum (ER) stress, redox stress, and nutrient deprivation. The functional role of CHOP10 in apoptosis and inhibition of adipose differentiation has already been reported.
One of the factors affecting fat metabolism is exercise. Exercise programs play a significant role in modulating the levels of adipokines secreted from adipose tissue and insulin resistance by reducing the number of fat cells, improving the secretory function of these cells, and reducing the content of macrophages in adipose tissue. It has been shown that the type and intensity of physical activity have different effects on energy metabolism. Aerobic exercises such as walking, cycling are low-cost therapeutic methods that are performed at low to moderate intensity for at least 30 minutes and 5 sessions per week to reduce metabolic risks, although, people are not able to implement these programs regularly. Therefore, there is a need for different intensities of aerobic exercises. High-intensity aerobic exercise has beneficial effects on important metabolic risk factors compared to low-intensity aerobic exercise. According to studies, adding intensity to exercises is more effective than adding volume to exercises in improving metabolic diseases. In recent years, there have been extensive studies on high-intensity training (HIT) and high-volume training (HVT) in terms of their differences and similarities in adaptations. The present study aims to investigate the effect of 8 weeks of HIT compared to HVT on expression of TCF4 and CHOP10 genes in subcutaneous adipose tissue in male rats.
Methods
In this experimental study, 32 male Wistar rats with an mean age of 6 weeks and a mean weight of 237±33 g were selected. Then, they were randomly divided into four groups of 8 including control group, moderate-intensity training (MIT), high-intensity training (HIT), high-intensity interval training (HIIT). The MIT group ran at a constant intensity of 65% of VO2max for 47 minutes. The HIT group ran at a speed of 20 meters per minute with an increasing slope for 40 minutes, and the HIIT group trained at an intensity of 90-100% of VO2max for 37 minutes. Adipose tissue sampling was done 48 hours after the last training session to measure the expressions of TCF4 and CHOP10 genes by real-time PCR method. After collecting the data, they were analyzed in SPSS software, version 20 statistical. The significance level was P<0.05. First, the normality of data distribution was examined using Shapiro-Wilk test. To determine the significance of the difference in variables among the study groups, Kruskal-Wallis test was used followed by the least significant difference test.
Results
The results of the Kruskal-Wallis test showed a significant difference between the three groups of HIIT, MIT and HIT in expression of TCF4 (P=0.011) and CHOP10 (P=0.008) genes (Figures 1 and 2) in subcutaneous adipose tissue of male Wistar rats.

There was a significant difference between the HIIT and control groups in TCF4 (P=0.006) and CHOP10 (P=0.004), but no significant difference was observed between the other groups.

Discussion
Overexpression of TCF4 causes higher production of primary adipocytes and their differentiation into subcutaneous fat. If this gene is overexpressed, it will cause obesity and related diseases such as hyperlipidemia, heart diseases, and diabetes. Overexpression of TCF4, which disrupts Wnt signaling pathway, stimulates the differentiation of primary adipocytes into mature adipocytes. Our results are consistent with the results of Fayaz et al. (2019), who showed that moderate-intensity aerobic activity reduced Wnt signaling compared to HIIT. Markewicz et al. (2019) concluded that TCF4 causes the differentiation of adipose tissue.
Evidence shows that CHOP10 temporarily inhibit the growth of primary adipocytes. Based on the findings of our study, HIIT increased the expression of CHOP10; as a result, primary fat cells were transformed into mature fat cells. Previous studies have shown that CHOP10 can protect cell survival by the regulation of autophagy in the early stages (before irreversible ER stress). Our results are consistent with the results of Bozi et al. (2016), who showed that aerobic exercise increased the amount of CHOP protein in a rate model of heart failure. In another study, it was reported that exercise increased the expression of ER proteins in the liver of obese rats and improved mitochondrial biogenesis.
More studies are needed to accurately understand the effective mechanisms caused by exercise. Since the role of exercise on mitochondrial biogenesis and fat metabolism has already been identified in many studies and it has been reported that the PGC1α-ATF6α signaling pathway increases CHOP protein level in skeletal muscles, more study on this signaling pathway that is known as ER homeostasis, is recommended in skeletal or cardiac muscles.
It can be concluded that the expression of TCF4 and CHOP10 decreased after HIIT. These findings show that HIIT is effective in controlling the expression of genes involved in fat metabolism.
Ethical Considerations
Compliance with ethical guidelines
The procedures for conducting this study were registered in the ethics committee of Payam Noor University with the number IR.PNU.REC.1399.083. It should be noted that the animals in this study were treated in accordance with the guidelines for care and work with laboratory animals of Helsinki 1964.
Funding
This article is based on Melika Khoshbeen's master's degree thesis, which was registered in the Department of Physical Education and Sports Sciences of Payam Noor Alborz University. All research costs have been borne by the researcher.
Authors' contributions
All authors contributed equally in preparing all parts of the research.
Conflict of interest
The authors declared no conflict of interest.
Acknowledgments
We are grateful to the laboratory officials of Payam Noor Alborz University who cooperated with us in this research.
مقدمه
بافت چربی نوعی از بافت پیوندی است که نقش فیزیولوژیکی مهمی را در پستانداران ایفا میکند. در ابتدا تصور میشد بافت چربی صرفاً بهعنوان یک انبار ذخیره چربی عمل میکند، اما امروزه بافت چربی بهعنوان یک اندام اندوکرین فعال شناخته شده است که مولکولهای بیواکتیو را که آدیپوکاین نامیده میشوند، ترشح میکند که در تنظیم هموستاز متابولیک مشارکت میکنند [1]. برخی از تحقیقات ثابت کردهاند علاوهبر تغذیه، برخی از ژنها نیز در رشد و تکثیر بافت چربی زیرجلدی تأثیر دارند [2]. یکی از این ژنها، پروتئین فاکتور رونویسی4 است. پروتئین فاکتور رونویسی4 بهعنوان فاکتور رونویسی عمل میکند که به نقوش تقویتکننده ایمونوگلوبولین mu-E5/kappa-E2 متصل میشود. فاکتور رونویسی4، رونویسی را با اتصال به (CANNTG-3-5’) E که معمولاً در SSTR2-INR یا عنصر آغازگر گیرنده سوماتوستاتین2 یافت میشود، فعال میکند [3، 4]. فاکتور رونویسی 4 در درجه اول با ایجاد تمایز عصبی با اتصال به دیاناِی در رشد عصبی جنین در دوران بارداری نقش دارد، درحالیکه در بزرگسالی فاکتور رونویسی4 در لنفوسیتها، عضلات اسکلتی و دستگاه گوارش یافت میشود [5].
جهش در فاکتور رونویسی4 باعث مشکلات گوارشی میشود که ممکن است نتیجه آن رشد و تکثیر سلولهای چربی زیرپوستی باشد [6]. اما یافتههای دیگری نیز نشان میدهد برخی از ژنها نیز بهعنوان آنتیفاکتور رونویسی 4 عمل کرده و مانع از تکثیر و رونویسی این ژن میشوند. یکی از این ژنها پروتئین همولوگ است که بهعنوان مهارکننده عماکرد این ژن عمل میکند [7]. پروتئین همولوگ یک پروتئین bZIP است که در تمایز و آپوپتوز نقش دارد. پروتئین همولوگ در انواع مسیرهای پاسخ به استرس، ازجمله استرس شبکه آندوپلاسمی (ER)، فشار ردوکس و محرومیت از مواد مغذی نقش دارد [8]. نقش عملکردی پروتئین همولوگC/EBP در آپوپتوز [9] و مهار تمایز چربی [10] گزارش شده است. با ترویج تخریب سلولهای β در پاتوژنز دیابت نقش دارد [11]. بیان پروتئین همولوگC/EBP را میتوان با استفاده از سایتوکاینهای التهابی ازطریق سیگنالینگ نیتریک اکساید ایجاد کرد [12]. اختلال در ژنپروتئین همولوگC/EBP شروع دیابت ناشی از استرس شبکه آندوپلاسمی را کند میکند و این نشان میدهد همولوگC/EBP فنوتیپ دیابتی را تشدید میکند [11].
عواملی ازقبیل افزایش میزان مصرف چربی و افزایش بیان ژنی سایتوکاینها، به افزایش فراخوانی ماکروفاژها به بافت چربی زیرجلدی منجر میشوند [13]. یکی از عوامل تأثیرگذار بر متابولیسم چربی، انجام تمرینات ورزشی است. برنامههای تمرینی با کاهش در تعداد سلولهای چربی، بهبود عملکرد ترشحی این سلولها و کاهش محتوای ماکروفاژهای بافت چربی، در تعدیل سطوح آدیپوکاینهای ترشحشده از بافت چربی و مقاومت انسولینی نقش بسزایی دارد [14]. مطالعات نشان دادند تمرین و فعالیت بدنی، میزان اکسیداسیون کل بدن را افزایش می دهد و از این طریق انرژی لازم برای عضلات را فراهم میکند و نیمرخ لیپید را بهبود میبخشد [15]. هرچند برخی از مطالعات، عدم تغییر نیمرخ لیپید به دنبال 8 هفته تمرین هوازی را گزارش کردهاند [16].
علاوهبر موارد یادشده عدم فعالیت بدنی نیز یکی از فاکتور اصلی مرتبط با چاقی است [17]. تمرین ورزشی آنزیمهای لیپوژنیک و بیان ژنهای دی نولیپوژنز مانند اسید چرب سنتتاز پروتئین پیوندیc1 تنظیمکننده استرول را کاهش میدهد و دسترسی به اسیدهای چرب زنجیره بلند برای سنتز تریگلیسرید را تنظیم میکند [14]. نشان داده شده است که نوع و شدت فعالیت بدنی اثرات متفاوتی را بر متابولیسم انرژی ایفا میکنند [18]. مطالعات فراوانی تأثیر تمرین هوازی را بر چاقی و کاهش وزن بررسی کردهاند [19، 20]. فعالیتهای ورزشی هوازی مانند پیادهروی، دوچرخهسواری صرفنظر از مکان، روشهای کمهزینه درمانی هستند که با شدت پایین تا متوسط با زمان حداقل 30 دقیقه و 5 جلسه در هفته بهمنظور کاهش خطرات متابولیکی انجام میشوند. هرچند، اغلب مردم بهطور منظم قادر نیستند این برنامهها را اجرا کنند [21]. ازاینرو نیاز به شدتهای مختلف تمرین ورزشی مناسب است.
تمرینات هوازی شدید در مقایسه با تمرینات هوازی با شدت کم مزایای سودمندی را بر فاکتورهای خطرزای مهم متابولیسمی دارند [22] . همچنین بنابر مطالعات انجامشده، شدت فعالیت ورزشی در مقابل حجم فعالیت عاملی مؤثرتر بر بهبود بیماریهای متابولیکی است [23]. در سالهای گذشته بحثهای گستردهای درباره تمرین شدید و تمرینات با حجم بالا ازحیث تفاوتها و شباهتهایشان در سازگاریها شکل گرفته است. نشان داده شده است تمرین شدید نهتنها یک ابزار مناسب و سودمند برای ورزشکاران نخبه، بلکه برای جلوگیری از آسیب و توانبخشی نیز مفید است. بیشتر پژوهشها بر تغییرات عملکرد استقامتی، سازگاریهای عضلانی یا فواید سلامتی، بهمنظور ایدههای بهتری درزمینه مکانیسمهای مولکولی، تمرکز کردهاند [21].
به نظر میرسد افزایش تخریب ژنوم و استرس اکسیداتیو در گسترش بافت چربی زیرپوستی بدن، بهدلیل سرعت بالای فرایندهای متابولیک جهت حفظ اعمال بیولوژیکی در این افراد باشد که میتواند به افزایش تولید رادیکالهای آزاد و تخریب ژنوم منجر شود [19].
مزیتهای فعالیت بدنی منظم در سلامتی بهخوبی اثبات شده است. برخی مطالعات نشان دادهاند تمرینات هوازی منظم با شدت متوسط، با کاهش تحریک سمپاتیکی و افزایش آدیپوسایتوکاینهای ضدالتهابی، میزان رهایش میانجیهای التهابیـکه در ابتلا به بیماریهای مزمن نقش مهمی دارندـ از بافت چربی را مهار میکنند [23].
برای به دست آوردن اطلاعات بیشتر درزمینه کنترل تعادل متابولیسم چربی، بهویژه به دنبال تمرینات با شدتهای مختلف که اطلاعات کمی در این رابطه در دست است، به مطالعه ژنها و مسیرهای متابولیک در شرایط مختلف نیاز است.
بنابراین هدف پژوهش حاضر بررسی تأثیر شدتهای مختلف تمرینات هوازی بر بیان ژنهای فاکتور رونویسی 4 و پروتئین همولوگ در بافت چربی زیرجلدی رتهای نر نژاد ویستار است. چهبسا نتایج این تحقیق گامی باشد در جهت برنامهریزی دقیقتر برای تمرینات ورزشی منظمتر در میان ورزشکاران و مربیان ورزشی رشتههای مختلف که نتایج آن در کل جامعه ورزشی اثربخش خواهد بود.
روش بررسی
پژوهش حاضر از نوع تجربی و بنیادی است و بر روی 32 سر موش نر ویستار 8 هفتهای با میانگین وزن بدن 33±237 گرم بهعنوان نمونه تحقیق (خریداریشده از انستیتو رازی) انجام شد. موشها در گروههای 8 تایی و در محیطی با میانگین دمای 1/4±22 درجه سانتیگراد، رطوبت 4±۵۵ درصد و چرخه روشناییتاریکی 12:12 ساعت در قفسهای مخصوص از جنس پلیکربنات نگهداری شدند. تمامی حیوانات به آب و غذای ویژه موش دسترسی آزاد داشتند. غذای آزمودنیهای این پژوهش، تولید شرکت خوراک دام بهپرور بود. در تمام مراحل پژوهش، آب موردنیاز حیوانات بهصورت آزاد در اختیار آنها قرارداده شد. تمامی مراحل نگهداری و کشتار موشها براساس دستورالعمل نگهداری حیوانات آزمایشگاهی انجام شد.
موشها بهصورت تصادفی ساده به 4 گروه 8 تایی تقسیم شدند: کنترل 8 هفته، تمرین هوازی با شدت متوسط، تمرین هوازی پرشدت و تمرین هوازی تناوبی پرشدت. همچنین در همین زمان موشها در گروههای تمرین و کنترل با تردمیل آشنا شدند. ابتدا 2 هفته و 5 جلسه در هر هفته آشناسازی رتها با تمرینات ورزشی 15 دقیقه با سرعت 5 تا 15 متر انجام شد. پس از تقسیمبندی رتها در گروههای تمرینی، توان هوازی میانگین رتهای هر گروه براساس پروتکل هویدال و همکاران ارزیابی شد و شدت تمرینی هفته اول هر گروه مشخص شد. تمرینات 8 هفته و 5 روز در هفته براساس دستورالعمل شروع شد. کلیه تمرینات در صبح و براساس ترتیب مشخص انجام شد. گروه کنترل 8 هفته همراه با 4 گروه تمرینی پس از 8 هفته تمرین و 24 ساعت پس از آخرین جلسه تمرینی و 12 ساعت ناشتایی بافتبرداری شدند. حیوانات با تزریق زایلازین و کتامین بیهوش شدند و برداشت بافت چربی زیرجلدی بلافاصله انجام شد. بافت نمونه هر حیوان بلافاصله در تیوب وارد محلول نیتروژن مایع شد و در دمای منفی 80 درجه فریز شد. نمونهها در آزمایشگاه تا زمان انجام آزمایشات ارزیابی مقدار تغییرات بیان ژن در فریزر منهای 80 درجه نگهداری شدند.
حداکثر اکسیژن مصرفی حیوانات باتوجهبه عدم دسترسی به ابزار مستقیم، با آزمون فزاینده بر روی نوارگردان مطابق با پروتکل هویدال و همکاران [1] و با پروتکل غیرمستقیم ارزیابی شد. ابتدا 10 دقیقه گرم کردن با شدت 40 تا 50 درصد VO2max انجام شد. سپس رتها با سرعت 15 متر بر دقیقه، 2 دقیقه شروع به دویدن کردند و هر 2 دقیقه یکبار به میزان 2 متر بر دقیقه تا سر حد واماندگی سرعت افزایش یافت. موشها 8 هفته پس از 5 دقیقه گرم کردن (با سرعت 5 متر بر دقیقه) به فعالیت پرداختند. تعداد جلسات در هر هفته 5 جلسه بود. پروتکل تمرین هوازی با شدت متوسط شامل دویدن در 65 درصد VO2max در زمان کل 47 دقیقه بود. تمرین شامل 5 دقیقه گرم کردن و 5 دقیقه سرد کردن و 37 دقیقه بدنه اصلی تمرین در 65 درصد اکسیژن مصرفی بیشینه بود. پروتکل تمرین هوازی پرشدت شامل دویدن در سرعت 20 متر بر دقیقه در زمان 40 دقیقه و با شیب فزاینده نوارگردان بود. تمرین شامل 5 دقیقه گرم کردن و 5 دقیقه سرد کردن و 30 دقیقه بدنه اصلی تمرین در 65 درصد اکسیژن مصرفی بیشینه بود.
شیب تردمیل در هفته اول صفر بود و هر 2 هفته، 2 درصد بر شیب افزوده شد تا در هفته هشتم به 8 درصد رسید [2]. پروتکل تمرین هوازی تناوبی پرشدت شامل 4 وهله تناوب شدید با زمان 4 دقیقه دویدن با شدت 90 تا 100 درصد VO2max و 4 وهله تناوب کمشدت با زمان 3 دقیقه دویدن در 50 تا 60 درصد VO2max بود که درمجموع 38 دقیقه طول کشید و شامل 10 دقیقه گرم کردن و 28 دقیقه بدنه اصلی تمرین بود. برای اطمینان از ایزولود بودن تمرین در هر 3 گروه تمرینات ورزشی براساس روش روکنمو و همکاران عمل شد. براساس این روش زمان خالص تمرین در هر گروه براساس زمان، شدت و تکرار وهلههای کار محاسبه و یکسان شد (فرمول شماره 1).

بنابراین با این روش مجموع 28 دقیقه تمرین تناوبی در شدتهای میانگین 95 و 55 درصد VO2max معادل 38 دقیقه تداومی در شدت 65 درصد VO2max محاسبه شد. بر همین منوال شدت تمرینات تداومی پرشدت نیز معادلسازی شد. بهمنظور از بین بردن اثرات حاد تمرین، 48 ساعت بعد از آخرین جلسه تمرینی، نمونهبرداری در هر گروه انجام شد. موشهای صحرایی با تزریق درونصفاقی زایلازین و کتامین بیهوش شدند و بافت کبد جدا و در سرم فیزیولوژیک شستوشو داده شد و سپس بلافاصله در میکروتیوب قرار داده شد و با استفاده از ازت مایع منجمد شد و برای سنجشهای بعدی به فریزر منهای 80 درجه انتقال داده شد.
بیان ژنهای TCF4 و CHOP10 با روش واکنش زنجیرهای پلیمراز در زمان واقعی بررسی شد. استخراج ریبونوکلئیک اسید بهصورت RNX-Pluse انجام شد و بهمنظور بررسی کیفیت و کمیت آن از روش اسپکتروفومتری و الکتروفورز بروی ژل آگارز استفاده شد. توالی mRNA مربوط به ژنهای TCF4 و CHOP10 با استفاده از مرکز ملی اطلاعات بیوتکنولوژی استخراج شد. پرایمرها توسط نرمافزار Allel ID ساخته شد و سپس هر پرایمر با نرمافزار BLAST جهت اطمینان از یکتا بودن محل جفت شدن پرایمرها ارزیابی شد (جدول شماره 1) (شرکت سینا ژن). آزمایشات جهت بررسی هر ژن، 2 بار انجام شد.

برای کمیسازی مقادیر بیان ژن موردنظر از فرمول ΔΔCT-2 استفاده شد. اطلاعات موردنیاز پس از جمعآوری، ازطریق نسخه 20 نرمافزار آماری SPSS در سطح معناداری 0/05>P پردازش و تحلیل شدند و کلیه نتایج بهصورت میانگین و انحرافمعیار بیان شدند. ابتدا طبیعی بودن توزیع دادهها با آزمون شاپیروویلک بررسی شد. برای تعیین معنادار بودن تفاوت بین متغیرها در 3 گروه یا بیشتر از آزمون آماری تحلیل واریانس (آنووا) یکراهه (کروسکال والیس) همراه با تست بونفرونی استفاده شد.
یافتهها
نتایج آزمون شاپیروویلک حاکی از طبیعی نبودن توزیع دادهها در هر گروه از تحقیق بود (0/05>P). همچنین یافتههای آنتروپومتریک نمونههای پژوهش حاضر در جدول شماره 2 نشان داده شده است.
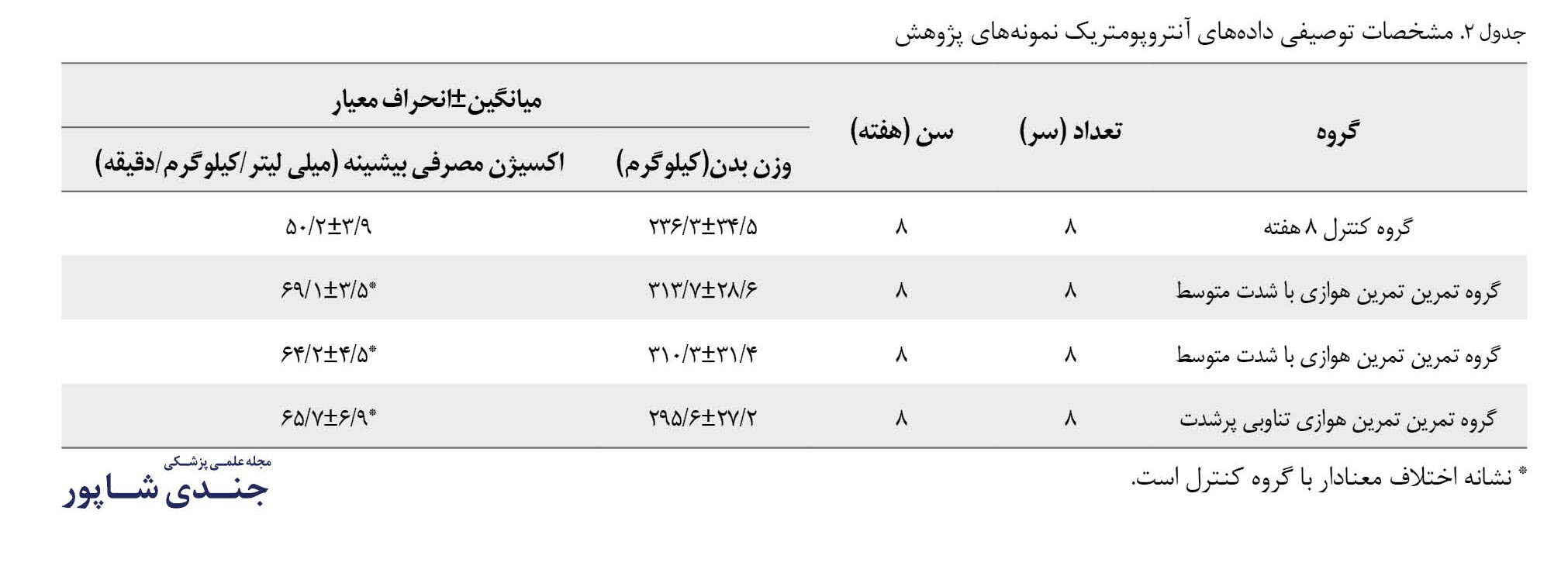
نتایج آزمون کروسکال والیس نشان داد بین اثر 3 شیوه تمرینات هوازی با شدت متوسط، تمرین هوازی پرشدت و تمرین هوازی تناوبی پرشدت بر بیان ژن فاکتور رونویسی 4 (تصویر شماره 1) و پروتئین همولوگ (تصویر شماره 2) بافت چربی زیرجلدی رتهای نر ویستار تفاوت معناداری وجود ندارد. نتایج بینگروهی نشان داد، بین گروه تمرین هوازی تناوبی پرشدت و کنترل تفاوت معناداری در بیان ژن فاکتور رونویس 4 (0/006=P) و پروتئین همولوگC/EBP -10 (P=0/004) بافت چربی زیرجلدی رتهای نر ویستار وجود دارد، اما بین دیگر گروهها تفاوت معناداری در مقادیر متغیرهای ذکرشده مشاهده نشد.


بحث
نتایج پژوهش حاضر نشان داد بین شدتهای مختلف تمرین هوازی با شدت متوسط، تمرین هوازی تناوبی پرشدت و تمرین هوازی پرشدت بر بیان ژن فاکتور رونویسی4 در بافت چربی رتهای نر ویستار در گروههای پژوهش تفاوت معناداری وجود ندارد. مقایسه بینگروهی نشان داد تفاوت معناداری در بیان ژن فاکتور رونویسی4 در بافت چربی زیرجلدی رتهای نر ویستار میان گروه تمرین هوازی تناوبی پرشدت نسبت به گروههای کنترل وجود دارد. بااینحال میان گروههای تمرین هوازی تناوبی پرشدت و تمرین هوازی پرشدت تفاوت معناداری مشاهده نشد. ضمناً اختلاف معناداری میان گروه تمرین هوازی با شدت متوسط و کنترل و تمرین هوازی پرشدت مشاهده نشد.
بررسی آزمون تعقیبی در گروههای تمرینی نشان داد اختلاف معناداری در بیان ژن فاکتور رونویسی4 میان گروههای کنترل و تمرین هوازی پرشدت وجود دارد. براساس یافتههای حاصل از پژوهش ما مشخص شد، تمرین هوازی تناوبی پرشدت بیان ژن فاکتور رونویسی4 را در بافت چربی زیرجلدی رتهای نر ویستار کاهش میدهد. با بررسی در نتایج بررسیهای صورتگرفته مشخص میشود که بیان بیشازحد فاکتور رونویسی4 باعث تولید بیشتر سلولهای چربی اولیه و تمایز آنها به چربی زیرجلدی میشود. درصورتیکه این ژن بیشازحد بیان شود باعث به وجود آمدن چاقی و بیماریهای مرتبط با آن مانند چربی خون، بیماریهای قلبی و دیابت میشود [24].
بیان بیش از حد فاکتور رونویسی 4 منفی غالب که سیگنالینگ Wnt را مختل میکند، تمایز سلولهای چربی اولیه به سلولهای چربی بالغ را تحریک میکند. درحالیکه فعالسازی سیگنالینگ Wnt ازطریق فاکتور رونویسی 4 توسط کلرید لیتیم باعث تمایز سلولهای چربی میشود [25]. اتصال Wnts مختلف به گیرندههای جزءبهجزء مربوطه و پروتئینهای مربوط به گیرندههای لیپوپروتئین با چگالی کم مسیرهای سیگنالینگ را فعال میکند که بیان ژن و عملکرد سلول را تغییر میدهد. مسیر Wnt متعارف به بسیج β-کاتنین در هسته منجر میشود. در آنجا خانواده فاکتورهای رونویسی TCF / LEF را فعال میکند. قرار گرفتن سلولهای پیشساز سلول چربی در معرض Wnts یا بیان خارجرحمی یک فرم فعال سازنده β- کاتنین با جلوگیری از القای PPARγ و C / EBPα مانع از چربیزایی میشود [26-28]. مکانیسم دقیق درگیر شناخته شده نیست، اما احتمالاً شامل بیان ژنهای TCF/LEF است زیرا بیان TCF غالب منفی (dnTCF) تا حدی اثرات مهاری Wnt را نجات میدهد [27، 28]. علاوهبراین، بیان dnTCF باعث تمایز خودبهخود سلولهای پیش سلول میشود که نشان میدهد مسیر سیگنالینگ Wnt متعارف در سلولهای پیشساز برای سرکوب چربیسازی عمل میکند.
یک کاندید جذاب برای یک مهارکننده چربیزایی ناشی از TCF، سیکلین D1 است؛ زیرا ژن آن یک هدف مستقیم از سیگنالینگ Wnt است و نشان داده شده است که فعالیت PPARγ را متضاد میکند [29]. هنگامی که فاکتور رونویسی 4 با پروتئین E دیگری (TCF3 یا TCF12) یا با یک نسخه دیگر از خود همکاری میکند، تصور میشود دیمر پروتئین E مستقیماً رونویسی ژن محلی را در یک پروتئین جعبه-E هدف فعال کند. بااینحال، سایر شرکای اتصالدهنده bHLH ممکن است سرکوبگرهای رونویسی، مانند ETO یا mSin3A را در همان مکان ژنومی جذب کنند [27-29]. از طرف دیگر، فعالیت فاکتور رونویسی4 با مهارکننده پروتئینهای اتصالدهنده دیاناِی که خانوادهای از 4 پروتئین دامنه اصلی مارپیچ-حلقه کوتاه هستند که فاکتور رونویسی 4 را دنبال میکنند، مسدود میشود و این مانع از ارتباط دیاناِی میشود [30].
همراستا با پژوهش حاضر میتوان به نتایج یافتههای فیاض و همکاران [31] اشاره کرد که نشان دادند فعالیت هوازی با شدت متوسط در مقایسه با تمرین هوازی تناوبی پرشدت، سیگنالینگ Wnt را کاهش میدهد. اما در مقابل میتوان به تحقیق مارکویکز و همکاران [30] اشاره کرد که به این نتیجه رسیدند که فاکتور رونویسی4 باعث تمایز بافت چربی میشود. همچنین نتایج بررسی آنها نشان داد فاکتور رونویسی 4 در تمایز سلولهای دندریتیک پلاسماسیتوئیدی که بخشی از سیستم ایمنی سازگار را تشکیل میدهند، دخیل است. همچنین بیان ژن فاکتور رونویسی4 برای رشد سلولهای سازنده لنفوئید بسیار حیاتی است و باعث ایجاد سلولهای T و B میشود.
علاوهبراین، فاکتور رونویسی4 رشد سلولهای سرتولی، میوژنز، ملانوژنز و انتقال اپیتلیال مزانشیمی را تنظیم میکند. پس میتوان نتیجه گرفت افزایش بیان ژن فاکتور رونویسی4 ضمن اینکه باعث تبدیل چربیهای نوع اولیه به چربیهای بالغ میشود، میتواند باعث بهبود عملکرد سیستم ایمنی بدن شود. باوجوداین، این تنها مطالعهای است که تاکنون تأثیر یک مداخله فیزیکی (فعالیت ورزشی با شدتهای مختلف) را بر این ژن و مسیر سیگنالینگ آن را بررسی کرده است و برای مشخص شدن تأثیر تغییرات احتمالی ناشی از فعالیت بدنی بر آن، به تحقیقات گستردهتری نیاز است.
یافتههای پژوهش جاضر نشان داد بین 3 شیوه تمرین هوازی با شدت متوسط، تمرین هوازی پرشدت و تمرین هوازی تناوبی پرشدت بر بیان ژن پروتئین همولوگ C/EBP -10 در بافت چربی رتهای نر ویستار در گروههای پژوهش تفاوت معناداری وجود ندارد. مقایسه بینگروهی نشان داد تفاوت معناداری در بیان ژن پروتئین همولوگ C/EBP -10 در بافت چربی زیرجلدی رتهای نر ویستار میان گروه تمرین هوازی تناوبی پرشدت نسبت به گروههای تمرین هوازی با شدت متوسط و کنترل وجود دارد. بااینحال میان گروه های تمرین هوازی تناوبی پرشدت و تمرین هوازی پرشدت تفاوت معناداری مشاهده نشد. ضمناً اختلاف معناداری میان گروه تمرین هوازی با شدت متوسط و گروههای کنترل و تمرین هوازی پرشدت مشاهده شد. از سویی دیگر تفاوت معناداری میان گروه کنترل نسبت به گروه تمرین هوازی پرشدت مشاهده شد. براساس یافتههای حاصل از پژوهش حاضر مشخص شد. تمرین هوازی تناوبی پرشدت بیان ژن پروتئین همولوگ را در بافت چربی زیرجلدی رتهای نر ویستار کاهش میدهد.
بررسی نتایج نشان داد القای هورمون از سلولهای چربی اولیه رشد مرتبط باعث گسترش کلون میتوزی و به دنبال آن بیان پروتئین اتصالدهنده ژنهای CCAAT، و و تمایز در سلولهای چربی میشود. ترتیب این وقایع بسیار مهم است، زیرا C/EB ضدباکتری است و بهطور زودرس باعث گسترش کلونال میتوزی موردنیاز برای تمایز میشود.
C/EB یک فعالکننده رونویسی از ژن C/EB است و در اوایل برنامه تمایز بیان میشود، اما فاقد فعالیت اتصال به دیاناِی است و در محلیسازی به سانترومرها موفق نمیشود تا اینکه سلولهای چربی اولیه از پایانه G1-S از مسیر کلون میتوزی عبور کنند. شواهدی ارائه شده است که ژن پروتئین همولوگ رشد سلولهای چربی اولیه را بهطور موقت متوقف میکند. با رسیدن سلولهای چربی اولیه به فازS ، ژن پروتئین همولوگ با تنظیم کمتری همراه است، ظاهراً C/EB را از محدودیت مهاری رها میکند و امکان انتقال مجدد ژن C/EB را فراهم میکند. همچنین، ژن پروتئین همولوگ با تنظیم مهارکننده پروتئاز N-استیل-ال-لو-نورو، از فعال شدن C/EB ، بیان C/EB و چربی جلوگیری میکند [32، 33].
نتایج بررسی دادههای تحقیقات نشان میدهد پروتئین همولوگ C/EBP -10 که ازطریق تنظیم اتوفاژی در مراحل اولیه (قبل از استرس استرس شبکه آندوپلاسمی برگشتناپذیر) میتواند برای بقای سلولی محافظت کند. این فاکتور رونویسی پاسخگو به استرس بهطور گستردهای بهعنوان ارتباط بین استرس پروتئین تاشو طولانی، یعنی استرس شبکه آندوپلاسمی و آپوپتوز شناخته شده است [34]. باتوجهبه دانش ما، هنوز هیچ مطالعاتی نشان ندادهاند که پروتئین همولوگ C/EBP -10 مستقیماً به آپوپتوز منجر میشود. همراستا با نتایج پژوهش ما میتوان به نتایج تحقیق بوزی و همکاران [35] اشاره کرد که نشان دادند تمرینات هوازی سبب افزایش میزان پروتئین همولوگ C/EBP -10 در رتهای مدل سکته قلبی میشود. در پژوهش دیگری نیز گزارش شد که تمرین ورزشی بیان پروتئینهای شبکه اندوپلاسمی کبد موشهای چاق را افزایش میدهد و سبب بهبود بیوژنز میتوکندری میشود. باتوجهبه کمبود مطالعات درزمینه استرس شبکه اندوپلاسمی و بهخصوص ژن پروتئین همولوگ و تمرین ورزشی برای شناخت دقیق مکانیسمهای تأثیرگذار ناشی از فعالیت ورزشی به تحقیقات بیشتری نیاز است.
نتیجهگیری
نتایج پژوهش نشان داد میزان فاکتور رونویسی4 کاهش و پروتئین همولوگ ما نسبت به تمرینات ورزشی افزایش مییابد. این یافتهها نشان میدهند فعالیت ورزشی بهمنظور کنترل میزان بیان ژنهای دخیل در متابولیسم چربی مؤثر است.
ازآنجاییکه نقش تمرین ورزشی بر بیوژنز میتوکندری و متابولیسم چربی در تحقیقات بسیاری مشخص شده و از طرفی گزارش شده است که مسیر سیگنالینگ PGC1-α-ATF6α باعث افزایش پروتئین همولوگ C/EBP -10 عضله اسکلتی میشود [32]، پیشنهاد میشود در تحقیقی مشابه این مسیر سیگنالینگ که تحت عنوان هموستاز شبکه آندوپلاسمی شناخته میشود، در عضله اسکلتی یا قلبی بررسی شود.
یکی از مهمترین محدودیتها، تفاوتهای ژنتیکی رتها و تغییراتی است که ممکن است در مارکرهای مختلف در اثر افزایش سن در طول تحقیق ایجاد شده باشد و بر نتایج پژوهش مؤثر باشد.
محدودیتهای دیگر شامل عدم بررسی تغییرات متابولیکی در طی دوره تمرین و پس از آن و عدم همکاری صددرصدی حیوانات موردمطالعه در تمرینات ورزشی موردنظر محقق بود.
ملاحظات اخلاقی
پیروی از اصول اخلاق پژوهش
مراحل انجام این مطالعه در کمیته اخلاق دانشگاه پیام نور با شماره IR.PNU.REC.1399.083 ثبت شد. قابل ذکر است که از حیوانات در این مطالعه مطابق با دستورالعمل مراقبت و کار با حیوانات آزمایشگاهی هلسینکی 1964 تبعیت شد.
حامی مالی
این مقاله بر اساس پایاننامه مقطع کارشناسی ارشد ملیکا خوشبین که در گروه تربیت بدنی و علوم ورزشی دانشگاه پیام نور البرز ثبت شده است، اجرا شده است. تمامی هزینه های پژوهش بر عهده محقق بوده است.
مشارکت نویسندگان
تمام نویسندگان در آمادهسازی این مقاله مشارکت داشتهاند.
تعارض منافع
بنابر اظهار نویسندگان، این مقاله تعارض منافع ندارد.
تشکر و قدردانی
از مسئولین آزمایشگاه دانشگاه پیام نور البرز که در این پژوهش با ما همکاری کردند، قدردانی میشود.
References
